Wetenschap
Waarom zien mitochondriën eruit zoals ze eruit zien?

Krediet:Wikipedia commons
Een van de grootste uitdagingen in de hedendaagse biologie is het verklaren van de structuur van cristae, de binnenmembranen van mitochondriën. Een verklaring in dit geval is een reeks principes om te voorspellen welke vorm de cristae zal aannemen na basale metabolische manipulaties van de omgeving waarin de mitochondriën zich bevinden. Deze principes zullen daarom een beschrijving zijn van de ware functie van mitochondriën, iets dat tot nu toe alleen nauwelijks gedacht.
Recente ontwikkelingen in technieken zoals superresolutie-lichtmicroscopie met levende cellen en elektronentomografie hebben nieuw inzicht gegeven in het dynamische gedrag van cristae. Een gedetailleerde architectuur van het gehele mitochondriale volume kan nu worden opgebouwd uit een reeks kantelbeelden die worden teruggeprojecteerd om 3D-tomogrammen te creëren. Op maandag bespraken we hoe cristae worden veranderd en opnieuw geconfigureerd volgens de overvloed en gezondheid van verschillende binnenmembraan- en matrixeiwitten. Het voorwendsel voor die analyse waren de structurele en biochemische overeenkomsten tussen membranen in mitochondriën, thylakoïden en myeline die vermoedelijk helpen metabolieten te kanaliseren bij de productie van energie.
In een recent artikel van The Royal Society's Open Biology , verklaren onderzoekers de biogenese van cristae door de gecoördineerde activiteiten van vier belangrijke routes die evolutionair bewaard zijn gebleven, gaande van protisten en gisten tot hogere eukaryoten zoals wij:dimeervorming en oligomerisatie van het ATP-synthase op cristae-randen, assemblage van de 'mitochondriale contactplaats' en cristae organiserend systeem' (MICOS) op crista-juncties, membraanremodellering door een binnenmembraan-geassocieerd, dynamin-gerelateerd GTPase (Mgm1 in gist en OPA1 bij zoogdieren) en juiste aanpassing van de membraanlipidesamenstelling.
Voor de eerste route waarbij het ATP-synthase betrokken is, zijn verschillende dingen duidelijk. Zoals we eerder meldden, dicteert de spontane dimerisatie van ATP-synthase onder nauwkeurig gedefinieerde en soortafhankelijke hoeken in geordende rijen de geometrie op de begane grond. In tegenstelling tot de ademhalingscomplexen I-IV, die op het platte binnenste grensmembraan zijn geassembleerd, wordt ATP-synthase (complex V) volledig diep in de cristae-membranen geassembleerd. Hoewel verschillende ATP-synthase-subeenheid-eiwitten niet nodig zijn voor een goede cristae-vorming, zijn de Atp20- en Atp21-subeenheden strikt vereist.
Een overmaat aan ADP induceert een gecondenseerde conformatie met grote, gezwollen intra-kristalruimten. Daarentegen nemen mitochondriën onder ADP-beperkende omstandigheden de orthodoxe conformatie aan met gecontracteerde intra-kristalruimte. In de gigantische amoebe Chaos carolinensis bevatten mitochondriën normaal gesproken willekeurig georiënteerde buisvormige cristae. Bij verhongering nemen vergrote cristae een kubische morfologie aan met een zigzagachtig patroon. Bij muizen veroorzaken apoptotische middelen fusie van individuele cristae met daaropvolgende afgifte van cytochroom c uit de intrakristallijne ruimte in het grensgebied.
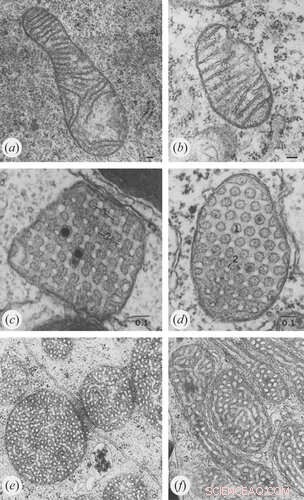
Credit:Klecker en Westermann, Open Biologie (2021). DOI:10.1098/rsob.210238
Voor de tweede route, de assemblage van MICOS-contactplaatsen, heeft onderzoek vastgesteld dat kritische eiwitten zoals die van de MIC60-gerelateerde genfamilie al aanwezig zijn in de endosymbiotische voorouders van mitochondriën - de α-proteobacteriën. Veel van deze eerdere mitochondriale voorouders vertonen al gedifferentieerde intracytoplasmatische membraanstructuren. Steevast missen soorten die hun mitochondriën zodanig hebben vereenvoudigd dat cristae afwezig zijn, de MICOS-gerelateerde genen. Herexpressie van MIC60-homologen in Δmic60-gistmutanten redt de ultrastructurele mitochondriale defecten.
De derde route omvat de aan dynamin gerelateerde GTPasen, die de fusie en splitsing van zowel binnen- als buitenmembranen coördineren. Bij splijting polymeriseren deze eiwitten tot samentrekkende ringen die samentrekkende krachten uitoefenen om mitochondriën samen te drukken. De uitkomst wordt nu geacht afhankelijk te zijn van interacties van deze eiwitten, zowel met het MICOS-complex en cristae-juncties, als ook met de transportsystemen van het binnen- en buitenmembraan die daar samenkomen. Deze omvatten de TIM- en TOM-membraantranslocatorcomplexen.
De vierde route omvat de mitochondriale membraanfosfolipiden zelf. Mitochondriën herbergen de biosyntheseroute van cardiolipine en zijn ook betrokken bij de synthese van fosfatidylethanolamine. Samen met fosfatidylcholine zijn dit de drie belangrijkste fosfolipiden waarmee mitochondriën werken. De meeste bouwstenen voor mitochondriale lipiden worden gesynthetiseerd in het ER en moeten daarom worden geïmporteerd door mechanismen die een nauwe aanhechting met het ER impliceren. Eenmaal in het buitenmembraan wordt de verdeling van lipiden gemedieerd door intermembraan-ruimte-gelokaliseerde transporteiwitten van de Ups/PRELI-familie,
Mitochondriën creëren geen geometrie vanuit het niets, maar benutten en bouwen eerder voort op de natuurlijke fysieke vormen die spontaan in lipiden voorkomen. Aan hun lot overgelaten, vormen lipiden concentrische lamellaire structuren die vervolgens kunnen worden uitgebreid en aangevuld met specifieke eiwitten. Fijnschalige metingen hebben nu aangetoond dat individuele cristae functioneel onafhankelijk zijn en significant verschillende membraanpotentialen kunnen hebben.
Cristae-formatie omvat een nauw verbonden samenspel van de bovengenoemde vier vormende invloeden. De activiteiten van het MICOS-complex en ATP-synthase-dimerisatie zijn bijvoorbeeld zowel coöperatief als antagonistisch. MICOS induceert negatieve membraankromming terwijl het ATP-synthase positieve kromming induceert bij cristae-uiteinden en -randen. Nieuwe computationele modellen, zoals momenteel in ontwikkeling in laboratoria over de hele wereld, waar de verhoudingen van deze verschillende componenten fijn kunnen worden aangepast en aangepast, zullen enorm helpen bij het definiëren van wat de vorm van mitochondriën regelt.
 Pig-Pen-effect:het mengen van huidolie en ozon kan een persoonlijke vervuilingswolk veroorzaken
Pig-Pen-effect:het mengen van huidolie en ozon kan een persoonlijke vervuilingswolk veroorzaken- Een nieuwe bindingsplaats voor antibiotica gevonden in het ribosoom
- Wetenschappers maken de kleinste onderling verbonden kettingen ter wereld
- Dominerende schimmel zou een oplossing kunnen zijn om meer biobrandstoffen en chemicaliën te produceren
- Antioxidanten en kunststoffen kunnen worden gemaakt van bijproducten van het malen van tarwe
- Nieuw onderzoek werpt licht op het effect van regen en wolken op atmosferische aerosolen
- Wetenschappers ontdekken dat seismische beeldvorming blind is voor water
- Explosieve uitbarsting doet vulkaan op Caribbeans St. Vincent
- George Monbiot Q + A - Hoe verjonging van de natuur de klimaatverandering kan helpen bestrijden
- Orkaan Nicole werpt licht op hoe stormen diepe oceaan beïnvloeden
Hoofdlijnen
- Gouden nanodeeltje gebruikt om virus te vervangen in nieuwe CRISPR-aanpak
- Twee nieuwe ontdekkingen werpen licht op het mysterie van hoe cellen omgaan met stress
- Het op lange termijn volgen van het voedselgedrag van walvissen via satelliet is nu mogelijk met nieuwe tag
- Hoe een 3D-dwarsdoorsnedemodel van de huid te maken
- Paranoia zal vernietigen:wanneer overschrijdt gezonde angst de grens in paranoia?
- What Is Crossing Over in Genetics?
- Cyanobacteriële studies onderzoeken de celstructuur tijdens stikstofgebrek
- Wetenschappers ontdekken een eeuwenoud geval van identiteitsverwisseling in de Chesapeake Bay
- Microalgen beloven overvloedig gezond voedsel en voer in elke omgeving
- Hoe kan een babyrobotzeehond een medisch hulpmiddel zijn?

- Hoe snel is een knipoog?
- Onverwachte bevinding in de energiecentrale van cellen
- Genetische barcodes worden gebruikt om cruciale populaties in een ecosysteem van koraalriffen te kwantificeren

- Biologische landbouw of bloemstroken:wat is beter voor bijen?

 Persoonlijke netwerken worden geassocieerd met het gebruik van schone brandstof voor het koken op het platteland van Zuid-India
Persoonlijke netwerken worden geassocieerd met het gebruik van schone brandstof voor het koken op het platteland van Zuid-India- Sociale media zijn niet allemaal slecht - het redt levens in rampgebieden
- Unieke studie omvat vloeistofdynamica en meer om te evalueren, toekomstige implantaten verbeteren
- NSA-lek voedt toename hacking voor cryptomining:rapport
- Nieuw lab-on-a-chip-platform wil de detectie van pathogenen verbeteren
- Beroemd economisch experiment dat duizenden keren is gereproduceerd
- 7 Classificaties van Earthworms
- Nanodragers kunnen nieuwe hoop bieden op therapie voor hersenkanker
- Elektronica
- Biologie
- Zonsverduistering
- Wiskunde
- French | Italian | Spanish | Portuguese | Swedish | German | Dutch | Danish | Norway |

-
Wetenschap © https://nl.scienceaq.com
