Wetenschap
Structureel bewijs voor een dynamische metallocofactor tijdens distikstofreductie door Mo-nitrogenase
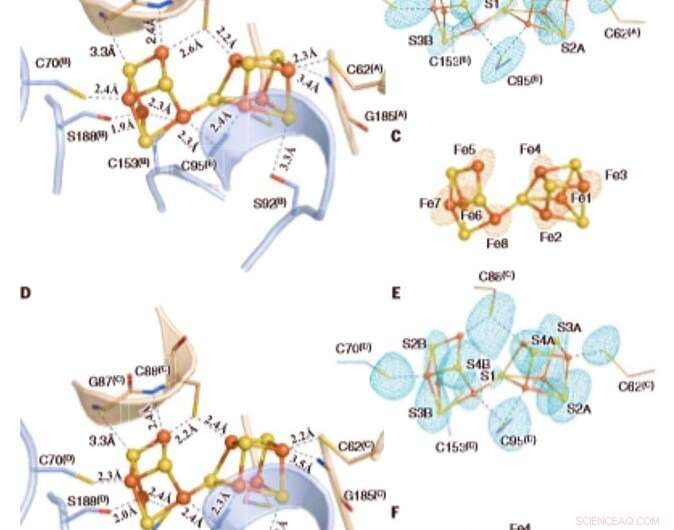
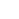
De geoxideerde P-clusters in Av1*. Structuren van P-clusters op (A tot C) het grensvlak van ketens A en B [P-cluster(A/B)] en (D tot F) het grensvlak van ketens C en D [P-cluster(C/D) ] van Av1*. Ketens A en C zijn de a-subeenheden, en ketens B en D zijn de b-subeenheden van de twee ab-dimeren van Av1*. [(A) en (D)] De P-clusters worden getoond in bal-en-stokpresentatie, en de sleutelresiduen die een interactie aangaan met de P-clusters worden aangegeven als stokjes. Kettingen A en C worden weergegeven als tarwelinten, en kettingen B en D worden weergegeven als lichtblauwe linten. [(B) en (C)] P-cluster(A/B) en [(E) en (F)] P-cluster(C/D) gesuperponeerd met [(B) en (E)] de afwijkende dichtheidskaarten berekend op 7100 eV met een resolutie van 2,18 Å en geprofileerd op 4,0 s, met de positie van zwavelatomen (mintblauw gaas); en met [(C) en (F)] de afwijkende dichtheidskaarten berekend op 7141 eV met een resolutie van 2,1 en geprofileerd op 15,0 s, met de positie van ijzeratomen (rood gaas). Atomen zijn als volgt gekleurd:Fe, Oranje; S, geel; O, rood; N, blauw. Afkortingen van één letter voor de aminozuurresiduen zijn als volgt:C, Cy; G, Gly; H, Zijn; R, arg; S, ser. Krediet:Wetenschap, doi:10.1126/science.aaz6748
Het enzym stikstofase is een biologische katalysator die distikstof (N2) kan reduceren tot ammoniak in aanwezigheid van een reeks complexe metallocofactoren. Echter, de mechanistische details van de reactie blijven schaars. In een nieuw verslag over Wetenschap , Wonchull Kang en een onderzoeksteam in de chemie, moleculaire biologie en biochemie aan de Universiteit van Californië-Irvine, ONS., rapporteerde een 1,83-angstrom-kristalstructuur voor het stikstofase-molybdeen-ijzer (MoFe) eiwit, die ze onder fysiologische distikstofomzetomstandigheden hebben gevangen. De resultaten van de studie kunnen de mogelijke mechanismen van N . beoordelen 2 reductie en de rol van gordelzwavelsites tijdens het proces.
Nitrogenase is een katalysator voor een cruciale stap in de wereldwijde stikstofcyclus, tijdens de omgevingsreductie van atmosferisch distikstof (N 2 ) aan de biologisch beschikbare ammoniak (NH 3 ). Het molybdeen-stikstofase-enzym bevat twee eiwitcomponenten:een met het ijzer (Fe) eiwit in een ijzerzwavel (Fe 4 S 4 ) cluster met een adenosinetrifosfaat (ATP)-bindingsplaats binnen elke subeenheid. De andere, molybdeen-ijzer (MoFe) eiwit met een α 2 β 2 heterotetrameer met twee complexe metalloclusters. Tijdens molybdeen-nitrogenase (Mo-nitrogenase) katalyse, de herhaalde associatie en dissociatie tussen de twee eiwitcomponenten maakte ATP-afhankelijke elektronenoverdracht van de Fe . mogelijk 4 S 4 cluster naar het MoFe-eiwit voor substraatreductie. Het vermogen van stikstofase om veel elektronen naar zijn cofactor te transporteren, maakte het enzym zeer veelzijdig tijdens substraatreductie.
Het werkingsmechanisme van het stikstofase-enzym begrijpen
Er zijn veel inspanningen geleverd om de mechanismen van stikstofase te begrijpen sinds de ontdekking ervan, waar sommigen zich hadden gericht op substraat- en remmerinteracties van het enzym. Van deze inspanningen Kang et al. een strategie bepaald die het overwegen waard is door het beperken van overtollige elektronenvoorraden die onbedoeld de N . aandreven 2 reductieproces naar voren. Dit bracht de substraat- of intermediaire toestand van het enzym terug naar een rusttoestand of reduceerde het enzym tot een niet waarneembare gemengde toestand. Het proces was relevant omdat stikstofase-eiwitten routinematig worden geïsoleerd in de aanwezigheid van een overmaat aan dithioniet als een extern geleverd reductiemiddel, en de verwijdering van deze kunstmatige elektronenbron in afwezigheid van zuurstof zou wetenschappers kunnen helpen distikstof (N 2 ) of zijn tussenproducten voor analyse.
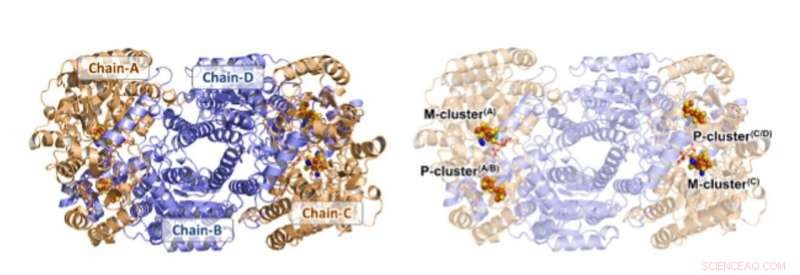
Structuur van Av1*. Solide (links) en transparante (rechter) lintpresentaties van de heterotetrameer van Av1*, met de a-subeenheden (Chain-A en Chain-C) en b-subeenheden (Chain-B en Chain-D) gekleurd in tarwe en blauw, respectievelijk. De M- en P-clusters zijn weergegeven als ruimtevullende modellen. Kleurcode van atomen:Fe, Oranje; S, geel; O, rood; N, blauw; ma, cyaan; C, grijs. De M-clusters in Chain-A en Chain-C worden aangeduid als M-cluster(A) en M-cluster(C), respectievelijk. De P-clusters op de interfaces Chain-A/Chain-B en Chain-C/Chain-D worden aangeduid als P-cluster(A/B) en P-cluster(C/D), respectievelijk. PYMOL werd gebruikt om dit cijfer voor te bereiden. Krediet:Wetenschap, doi:10.1126/science.aaz6748
Als proof-of-concept, Kang et al. bereidde het ruwe extract van een anerobe bacteriestam Azotobacter vinelandii met of zonder toevoeging van dithioniet na celverstoring. De A. vinelandii-stam bracht in beide gevallen actief een Mo-nitrogenase tot expressie die een met histidine gemerkt MoFe-eiwit bevatte. Toen ze de activiteit van deze monsters analyseerden, de dithionietvrije ruwe extractmonsters waren bijna inactief tijdens substraatreductie - vanwege uitputting van elektronen in de ruwe extracten tijdens celverstoring. Kang et al. zou daarom de activiteit van monsters volledig kunnen herstellen door dithioniet en ATP toe te voegen (d.w.z. door elektronen te leveren).
Clusters van stikstofase - twee unieke metalloclusters:de P-cluster en de M-cluster.
Op basis van de geschetste voorwaarden, wanneer een stikstofase tot expressie brengende cultuur die actief N . uitvoert 2 fixatie wordt onderworpen aan cellysis zonder extra elektronentoevoer, de stikstofase bleef functioneel. Hoewel mogelijk gestopt in een "slapende" of intermediaire toestand als gevolg van terugtrekking van de elektronenstroom in een ijzerzwavel-metallocluster die bekend staat als de M-cluster, gelokaliseerd in het stikstofase-enzym. Wanneer Kang et al. het dithionietvrije ruwe extract gezuiverd, het histidine-gelabelde MoFe-eiwit (aangeduid als AV1* in de studie) was actief tijdens N 2 reductie en ook volledig functioneel. Toen het team AV1* kristalliseerde, ze observeerden bruine kristallen die afbuigen tot een resolutie van 1,83 angström (Å). Ze bevestigden de structurele herschikking van de twee P-clusters van AV1 * met behulp van afwijkende dichtheidsgegevens en gebruikten paramagnetische elektronenresonantie om de structurele toewijzing te observeren. De resultaten gaven hen lang gezochte antwoorden op de fysiologische relevantie van deze experimentele toestand en wezen op een beperkte stroom van elektronen tussen de twee unieke metalloclusters (P- en M-clusters) van de verbinding in afwezigheid van dithioniet.
Een aannemelijk mechanisme van N2-reductie door Mo-nitrogenase. Krediet:Markus Ribbe en Yilin Hu, Wetenschap , doi:10.1126/science.aaz6748 
Een plausibel werkingsmechanisme dat overeenkwam met de experimentele waarnemingen omvatte de stapsgewijze vermindering van distikstof (N 2 ) op de drie banden zwavelplaatsen op de stikstofasekatalysator op basis van de rotatie van de M-cluster. Het voorgestelde mechanisme begint met een strakke binding van N 2 op een bepaalde plaats, gevolgd door rotatie van de gebonden N 2 naar een andere volgende plaats (plaatsen aangeduid als S3A tot S2B tot S5A op de verbinding). Tijdens het proces, de reductie/protonering van N 2 tot het diazeenniveau gebeurde door waterstofbinding, gevolgd door verdere reductie/protonering voor de omzetting in ammoniak, voordat het uit de structuur wordt losgelaten. Daaropvolgende rotatie van het cluster bracht een nieuwe N 2 molecuul naar de volgende plaats om de volgende ronde van stapsgewijze N . te starten 2 reductie door voortdurende clusterrotatie in een delicaat mechanisme tijdens katalyse. Dergelijke cycli tussen verschillende reactieplaatsen waren losjes analoog aan het mechanisme van het ATP-synthase-enzym. De roterende metallocluster maakte dus effectief de multi-elektronenreductie van N . mogelijk 2 via een verdeel-en-heers-aanpak.
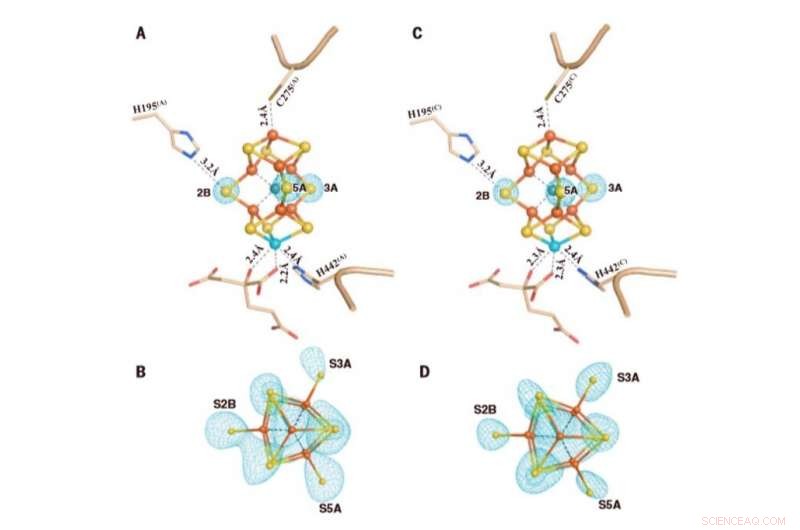
De M-clusters in Av1*(TOD). Structuren van M-clusters in (A en B) keten A [M-cluster(A)] en (C en D) keten C [M-cluster(C)] verfijnd met een resolutie van 1,73 Å. Side view of (A) M-cluster(A) and (C) M-cluster(C) with key residues interacting with the clusters indicated as sticks. M-cluster(A) and M-cluster(C) are superimposed with the Fo-Fc omit maps of the belt sulfurs contoured at 13s (mint-blue mesh). View along the Fe1-C-Mo direction of (B) M-cluster(A) and (D) M-cluster(C) superimposed with the anomalous density maps calculated at 7100 eV at a resolution of 2.17 Å and contoured at 4.0s, showing the presence of the anomalous sulfur density (mint-blue mesh) at all belt sulfur positions (S2B, S3A, and S5A) in (B) M-cluster(A) and (D) M-cluster(C). Credit:Science, doi:10.1126/science.aaz6748
To understand the sulfur-displaced conformation of AV1* under limited electron flux, the team formed AV1* turnover with dithionite (designated as AV1*TOD), to yield brown crystals that diffracted to a resolution of 1.73 Å. The observations were consistent with the mechanism of bound dinitrogen species on the compound and illustrated the physiological relevance of the conformation during catalysis. The capacity to displace three different sites by a dinitrogen species was consistent with previous investigations on catalysis-dependent selenium. Kang et al. proposed many mechanisms to explain the observations, however they seek further experimental support to verify them. The team highlighted the possibility for all belt-sulfur sites to be involved in the process of catalysis due to the presence of asymmetric belt-sulfur displacements in the compound. The results aim to provoke a paradigm shift in the mechanistic thinking of nitrogenase activity, ultimately to understand the intricate mechanism of the enzyme.
© 2020 Wetenschap X Netwerk
 Gebruik voor waterstofsulfide
Gebruik voor waterstofsulfide - Studie benadrukt belofte van galliumoxiden voor stralingsdetectoren van de volgende generatie
- Onderzoekers ontwikkelen verbeterd recyclingproces voor koolstofvezels
- Nieuw pathogeen mechanisme voor influenza NS1-eiwit gevonden
- Overlaat voor elektronen kan voorkomen dat lithium-metaalbatterijen vlam vatten
- Episodische en intense regen veroorzaakt door de oude opwarming van de aarde
- Tropische storm Faxai krijgt een naam en NASA krijgt een infraroodfoto
- Een deal voor de natuur om dieren in het wild te redden:WWF-chef
- Natuurlijke afzettingen van heliumgas kunnen helpen bij de veilige productie van schalie- of steenkoolgas, onderzoek suggereert:
- Het hardnekkige hogedruksysteem achter de recordhittegolven in Australië
Hoofdlijnen
- De genetische handtekening van de zeeschelp lezen
- Hoe chromosomen vals spelen om kans te maken in een ei te komen
- Een JELL-O-model van een diercel maken
- Meer dan 75 procent afname van de totale biomassa van vliegende insecten in 27 jaar
- Orgelsystemen betrokken bij homeostase
- Recombinante DNA-technologie voor vaccinontwikkeling
- Opwarming van de oceaan signaleert verandering van dieet voor Europese aalscholvers
- Structuur van het spierstelsel
- Hoe hoge bomen suikers verplaatsen
- Een nieuwe kijk op brillen

- Wetenschappers vinden mogelijke ontbrekende schakel in chemie die leidde tot leven op aarde

- Stabiliserende eigenschappen van vurengom kunnen worden aangepast met behulp van extractietechnieken

- Voordelen en nadelen van het gebruik van propaan

- Vast acetyleenreagens voor laboratoriumtoepassingen
 In vitro/in silico-evaluaties van bindingsaffiniteiten van perfluoralkylstoffen aan Baikal-afdichting PPARα
In vitro/in silico-evaluaties van bindingsaffiniteiten van perfluoralkylstoffen aan Baikal-afdichting PPARα- Mysterieuze uitbarsting kwam van Campi Flegrei caldera
- Onderzoekers ontwikkelen een procedure en een goedkope, snel en milieuvriendelijk apparaat dat bittere amandelen kan detecteren
- Groenlandse ijskap dreigt onomkeerbaar te smelten
- Hoe Grams in Milligrammen om te zetten
- Verband tussen het atoomnummer en de chemische reactiviteit van alkalimetalen
- Hoe ontvangt de aarde warmte van de zon?
- 3D-printen voor efficiënter chemisch onderzoek
- Elektronica
- Biologie
- Zonsverduistering
- Wiskunde
- French | Italian | Spanish | Portuguese | Swedish | German | Dutch | Danish | Norway |

-
Wetenschap © https://nl.scienceaq.com
